Ocean deoxygenation
Coastal hypoxia is an anthropogenic disturbance fueled by eutrophication that is accelerating worldwide. Low-oxygen areas are generally characterized by mass-mortality of marine life and resultant “dead zones” which can have significant impacts on ecosystem services. For example, we found that mortality and reduced growth of mussels in a hypoxic New England estuary led to a >75% drop in their ecosystem function of filtering bay waters. However, we also found that low oxygen conditions can enhance economically valuable shellfish by increasing their abundance due to disrupted food web interactions. Hypoxia causes mobile predators (e.g. fish and crabs) to flee stressful hypoxic areas, leaving sufficiently hypoxia-tolerant clams with a predation refuge. Combined with comparative physiological studies that revealed dramatic differences in the relative tolerance of several benthic species, our work suggests that a mechanistic understanding of organismal tolerances and species interactions is necessary for predicting ecosystem trajectories in response to hypoxia.
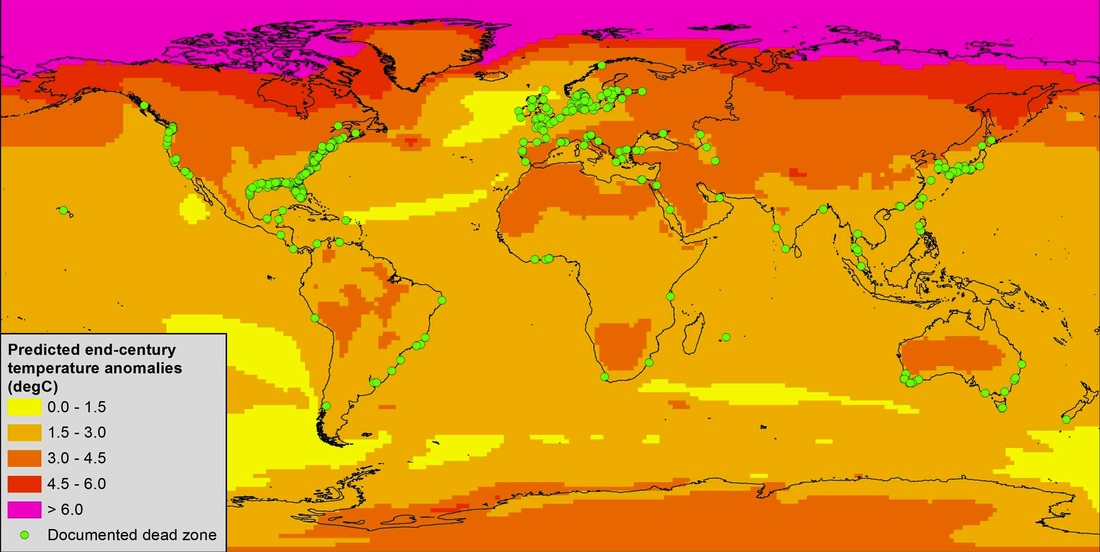
Our recent synthesis predicts that ocean deoxygenation will become more severe and prevalent with climate change because warming temperatures and other anticipated climate changes exacerbate hypoxia through mechanisms at organismal, community, and ecosystem levels. Moreover, we found that these impacts will occur widely since over 90% of known dead zones will experience a > 2°C temperature change by end of century. However, this is likely an underestimate of the dead zone threat since hypoxia has been studied almost exclusively in temperate regions. Our current work in the tropics suggests that dead zones can have catastrophic effects on coral reefs.
Coastal hypoxia is an anthropogenic disturbance fueled by eutrophication that is accelerating worldwide. Low-oxygen areas are generally characterized by mass-mortality of marine life and resultant “dead zones” which can have significant impacts on ecosystem services. For example, we found that mortality and reduced growth of mussels in a hypoxic New England estuary led to a >75% drop in their ecosystem function of filtering bay waters. However, we also found that low oxygen conditions can enhance economically valuable shellfish by increasing their abundance due to disrupted food web interactions. Hypoxia causes mobile predators (e.g. fish and crabs) to flee stressful hypoxic areas, leaving sufficiently hypoxia-tolerant clams with a predation refuge. Combined with comparative physiological studies that revealed dramatic differences in the relative tolerance of several benthic species, our work suggests that a mechanistic understanding of organismal tolerances and species interactions is necessary for predicting ecosystem trajectories in response to hypoxia.
Our recent synthesis predicts that ocean deoxygenation will become more severe and prevalent with climate change because warming temperatures and other anticipated climate changes exacerbate hypoxia through mechanisms at organismal, community, and ecosystem levels. Moreover, we found that these impacts will occur widely since over 90% of known dead zones will experience a > 2°C temperature change by end of century. However, this is likely an underestimate of the dead zone threat since hypoxia has been studied almost exclusively in temperate regions. Our current work in the tropics suggests that dead zones can have catastrophic effects on coral reefs.

|

|
Selected publications
Howard, R.D., M.D. Schul, L.M. Rodriguez Bravo, A.H. Altieri, J.L. Meyer (2023) Shifts in the coral microbiome in response to in situ experimental deoxygenation. Applied and Environmental Microbiology 89: e00577-23.
Mallon, J., A. Demko, J. Sneed, L. Newman, C. Dugan, A.H. Altieri, V.J. Paul, M.D. Johnson (2023) The influence of deoxygenation on Caribbean coral larval settlement and early survival. Frontiers in Marine Science 10: 1254965.
Johnson, M.D., S.D. Swaminathan, E.N. Nixon, V.J. Paul, A.H. Altieri (2021) Differential susceptibility of reef-building corals to deoxygenation reveals remarkable hypoxia tolerance. Scientific Reports 11: 1-12.
Johnson, M.D., J.J. Scott, M. Leray, N. Lucey, L.M.R. Bravo, W.L. Wied, A.H. Altieri (2021) Rapid ecosystem-scale consequences of acute deoxygenation on a Caribbean coral reef. Nature Communications 12: 1-12.
Altieri, A.H., M.D. Johnson, S.D. Swaminathan, H.R. Nelson, K.B. Gedan (2021) Resilience of tropical ecosystems to ocean deoxygenation. Trends in Ecology and Evolution 36: 227-238.
Altieri, A.H., H.R. Nelson, K.B. Gedan (2019) The significance of ocean deoxygenation for tropical ecosystems – corals, seagrasses, and mangroves. Pages 401-429 in Ocean Deoxygenation: Everyone’s problem – Causes, impacts, consequences and solutions. D, Loffoley & J.M. Baxter, editors. IUCN, Gland, Switzerland.
Nelson, H.R. and A.H. Altieri (2019) Oxygen: The universal currency on coral reefs. Coral Reefs: 38: 177-198
Altieri, A.H. and R.J. Diaz (2019) Dead zones: Oxygen depletion in coastal ecosystems. Pages 453-473 in World Seas: An Environmental Evaluation, Vol. III. C. Sheppard editor. Elsevier Academic Press, Oxford, UK.
Johnson M.D., L.M. Rodriguez, A.H. Altieri (2018) Shallow-water hypoxia and mass mortality on a Caribbean coral reef. Bulletin of Marine Science 94:143-144.
Altieri, A.H., S.B. Harrison, J. Seemann, R. Collin, R.J. Diaz, and N. Knowlton (2017) Tropical dead zones and mass mortalities on coral reefs. Proceedings of the National Academy of Sciences USA 114: 3660-3665.
Gedan, K.B., A.H. Altieri, I. Feller, R. Burrell, D. Breitburg (2017) Community composition in mangrove ponds with pulsed hypoxic and acidified conditions. Ecosphere 8: e02053.
Altieri, A.H. and K.B. Gedan (2015) Climate change and dead zones. Global Change Biology 21: 1395-1406.
Altieri, A.H. (2008) Dead zones enhance key fisheries species by providing predation refuge. Ecology 89: 2808-2818.
Altieri, A.H. (2006) Inducible variation in hypoxia tolerance across the intertidal-subtidal distribution of the blue mussel Mytilus edulis. Marine Ecology Progress Series 325: 295-300.
Altieri, A.H. and J.D. Witman (2006) Local extinction of a foundation species in a hypoxic estuary: integrating individuals to ecosystem. Ecology 87: 717-730.
Howard, R.D., M.D. Schul, L.M. Rodriguez Bravo, A.H. Altieri, J.L. Meyer (2023) Shifts in the coral microbiome in response to in situ experimental deoxygenation. Applied and Environmental Microbiology 89: e00577-23.
Mallon, J., A. Demko, J. Sneed, L. Newman, C. Dugan, A.H. Altieri, V.J. Paul, M.D. Johnson (2023) The influence of deoxygenation on Caribbean coral larval settlement and early survival. Frontiers in Marine Science 10: 1254965.
Johnson, M.D., S.D. Swaminathan, E.N. Nixon, V.J. Paul, A.H. Altieri (2021) Differential susceptibility of reef-building corals to deoxygenation reveals remarkable hypoxia tolerance. Scientific Reports 11: 1-12.
Johnson, M.D., J.J. Scott, M. Leray, N. Lucey, L.M.R. Bravo, W.L. Wied, A.H. Altieri (2021) Rapid ecosystem-scale consequences of acute deoxygenation on a Caribbean coral reef. Nature Communications 12: 1-12.
Altieri, A.H., M.D. Johnson, S.D. Swaminathan, H.R. Nelson, K.B. Gedan (2021) Resilience of tropical ecosystems to ocean deoxygenation. Trends in Ecology and Evolution 36: 227-238.
Altieri, A.H., H.R. Nelson, K.B. Gedan (2019) The significance of ocean deoxygenation for tropical ecosystems – corals, seagrasses, and mangroves. Pages 401-429 in Ocean Deoxygenation: Everyone’s problem – Causes, impacts, consequences and solutions. D, Loffoley & J.M. Baxter, editors. IUCN, Gland, Switzerland.
Nelson, H.R. and A.H. Altieri (2019) Oxygen: The universal currency on coral reefs. Coral Reefs: 38: 177-198
Altieri, A.H. and R.J. Diaz (2019) Dead zones: Oxygen depletion in coastal ecosystems. Pages 453-473 in World Seas: An Environmental Evaluation, Vol. III. C. Sheppard editor. Elsevier Academic Press, Oxford, UK.
Johnson M.D., L.M. Rodriguez, A.H. Altieri (2018) Shallow-water hypoxia and mass mortality on a Caribbean coral reef. Bulletin of Marine Science 94:143-144.
Altieri, A.H., S.B. Harrison, J. Seemann, R. Collin, R.J. Diaz, and N. Knowlton (2017) Tropical dead zones and mass mortalities on coral reefs. Proceedings of the National Academy of Sciences USA 114: 3660-3665.
Gedan, K.B., A.H. Altieri, I. Feller, R. Burrell, D. Breitburg (2017) Community composition in mangrove ponds with pulsed hypoxic and acidified conditions. Ecosphere 8: e02053.
Altieri, A.H. and K.B. Gedan (2015) Climate change and dead zones. Global Change Biology 21: 1395-1406.
Altieri, A.H. (2008) Dead zones enhance key fisheries species by providing predation refuge. Ecology 89: 2808-2818.
Altieri, A.H. (2006) Inducible variation in hypoxia tolerance across the intertidal-subtidal distribution of the blue mussel Mytilus edulis. Marine Ecology Progress Series 325: 295-300.
Altieri, A.H. and J.D. Witman (2006) Local extinction of a foundation species in a hypoxic estuary: integrating individuals to ecosystem. Ecology 87: 717-730.